
Die Gattung Pantherophis
Dies ist eine Übersicht über die diversen Änderungen innerhalb der Gattung Pantherophis.
Ausgangssituation | Die Kornnattern | Die Erdnattern | Die fuchsnattern | Quellen | Hinweis
Ausgangssituation
Die Gattung Pantherophis (Fitzinger, 1843) wurde von Utiger et al. (2002) im Rahmen einer umfassenden Analyse der Gattung Elaphe für die nordamerikanischen Kletternattern wieder eingeführt. Aktuell umfasst diese Gattung nun neun Arten, die sich grob in drei Gruppen einteilen lassen. Drei verschiedene Kornnattern, zwei Fuchsnattern und vier Erdnattern. Die Verbreitungsgebiete der Gruppen weisen größtenteils Überschneidungen auf, die einzelnen Arten in diesen Gruppen sind aber jeweils klar voneinander getrennt.
Die Kornnattern
Burbrink (2002) untersuchte die Verwandtschaftsverhältnisse der diversen Unterarten der Kornnattern (Elaphe guttata guttata, E. g. rosacea, E. g. emoryi, E. g. meahllmorum sowie E. g. intermontana). Da sich seine Arbeit mit der Umgruppierung durch Utiger et al. zeitlich überschnitt, war ihm der neue Gattungsname noch nicht bekannt. Die genetische Analyse zeigte, dass es sich um drei verschiedene geographische Linien handelt: Eine östliche -, eine zentrale - und eine westliche Linie, die durch geographische Barrieren oder unterschiedliche Lebensräume voneinander isoliert sind. Aufgrund der engen genetischen Ähnlichkeit von E. g. rosacea zu der Nominatform (E. g. guttata) der östlichen Linie wurde diese mit E. g. guttata synonymisiert. Die westliche Linie besteht aus E. g. emoryi an die sich südlich E. g. meahllmorum anschließt. Zwischen den beiden Unterarten gab es eine breite Gradationszone. Eine weitere geographisch isolierte Unterart, E. g. intermontana, war nicht allgemein anerkannt und ist genetisch beinahe identisch mit E. g. emoryi. Aus diesem Grund wurden beide mit E. g. emoryi synonymisiert. Jedoch ist eines der untersuchten Exemplare von E. g. meahllmorum aus Hidalgo, dem äußersten südöstlichen Rand von Texas näher mit Exemplaren aus der zentralen Linie verwandt und es ist möglich, dass es sich um eine weitere, bisher noch unbekannte Linie handelt. Die Tatsache, dass die östliche – und westliche Linie jeweils von einer einzigen Unterart dominiert sind (östliche Linie: E. g. guttata, westliche Linie: E. g. emoryi) brachte Burbrink (2002) dazu, beide bisherigen Unterarten in den Artstatus zu erheben. Für die zentrale Linie existierte bisher kein Art- oder Unterartname, so dass Burbrink für diese Art den Namen E. slowinskii vorschlug.
Im Jahr 2007 veröffentlichten Burbrink und Lawson (2007) eine Arbeit deren Ziel es war die Ausbreitung einiger nordamerikanischer Nattern (inklusive der Kornnattern) innerhalb der USA zu untersuchen und zeitlich einzuordnen. In dieser Arbeit verwendeten die Autoren unter anderem genetische Proben von Pantherophis guttatus, P. bairdi, P. obsoletus, P. vulpinus sowie Pituophis melanoleucus. Die Analyse zeigte, dass der einzige Vertreter der Kornnattern (P. guttatus) nicht so nahe mit den restlichen Arten verwandt war wie angenommen wurde. Allerdings wurde eine engere Verwandtschaft von Pituophis melanoleucus mit Pantherophis vulpinus festgestellt die es erforderlich machte eine Umgruppierung auf Gattungsebene vorzunehmen. Da der Name Pituophis (Holbrook, 1842) älter ist als Pantherophis (Fitzinger, 1843), wurden alle Arten der Gattung Pantherophis in die Gattung Pituophis überführt und Pantherophis mit dieser synonymisiert.
Allerdings waren die genetischen Analysen die zu dieser Änderung geführt haben aufgrund der geringen Anzahl an einbezogenen Arten und Exemplaren der jeweiligen Arten nicht sehr genau und aussagekräftig, so dass Collins und Taggart (2008) eine alternative Einteilung vorschlugen. Bei ihrer Einteilung berücksichtigten die Autoren zudem die morphologischen -, ökologischen - und biologischen Unterschiede der einzelnen Arten. Sie gliederten die drei Arten der Kornnatter (guttatus, emoryi, slowinskii) wieder in Pantherophis ein. Um den Konflikt den die nähere Verwandtschaft von Pituophis melanoleucus mit Pantherophis vulpinus verursachte und der die Umgruppierung von Pantherophis in Pituophis erforderlich gemacht hatte (s.o.) aufzulösen führten die Autoren zwei weitere Gattungen ein: Scotophis für die Erdnattern und Mintonius für die Fuchsnattern.
Daraufhin folgte eine erneute genetische Untersuchung von Pyron und Burbrink (2009). Diese Analyse war wesentlich detaillierter und genauer als die vorherige da diesmal alle betroffenen Arten mit einbezogen worden waren und bestätigte, dass die Einordnung von Pantherophis in Pituophis (Burbrink und Lawson, 2007) nicht die exakten Verwandtschaftsverhältnisse wiederspiegelte. Sie waren ebenfalls der Ansicht, dass auch die von Collins und Taggart (2008) vorgeschlagene Einteilung ungenau sei (siehe Abbildung und Erklärung). Demnach wurden die Kornnattern gemeinsam mit den Erd- und Fuchsnattern wieder der Gattung Pantherophis zugeschrieben.
Die Kritik an der Arbeit von Collins und Taggart (2008) bestand darin, dass diese Autoren die Verwandtschaftsverhältnisse unangemessen stark aufspalteten, indem sie neben Pantherophis und Pituophis noch zwei weitere Gattungen einführten (Scotophis für die Erdnattern und Mintonius für die Fuchsnattern), wodurch die eigentlich nahe genetische Verwandtschaft zwischen den Arten verschleiert wurde. Aus diesem Grund synonymisierten Pyron und Burbrink (2009) die neu eingeführten Gattungen mit Pantherophis.
Collins (2010) wies darauf hin, dass Pyron und Burbrinks (2009) Arbeit die von Collins und Taggart (2008) präsentierte Gattungseinteilung durchaus stütze und die Autoren keine Belege vorgebracht hätten, die diese Einteilung als falsch darstellt. Collins (2010) erwiderte die Kritik, die Pyron und Burbrink (2009) aufgebracht hatten und stellte die Frage, welche der beiden präsentierten Einteilungen die Verwandtschaftsverhältnisse eher verschleiern würde.
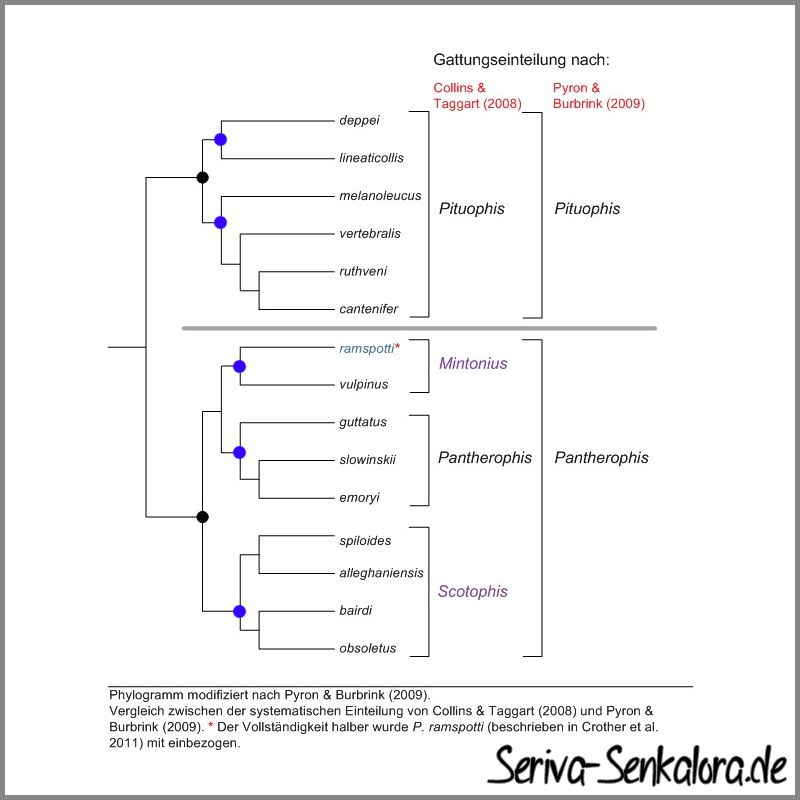
Anklicken zum Vergrößern
Um zu verdeutlichen, wieso sowohl Collins & Taggarts (2008) als auch Pyron & Burbrinks (2009) Ansatz richtig ist wurden beide Ansätze auf Basis eines Abstammungsbaums (Phylogramm) nebeneinander gestellt. Das Phylogramm zeigt die Abstammungsverhältnisse der Arten. Von den Artnamen ausgehend ist das Diagramm rückwärts zu lesen. Die Zusammenführung der Linien an denen die Artnamen hängen deutet einen gemeinsamen Vorfahren dieser Arten an (blaue Punkte). Man bezeichnet eine solche Gruppe von Arten als monophyletische Gruppe, also als Gruppe mit einem gemeinsamen Vorfahren. Dieses ist das Hauptkriterium zur Einteilung in eine eigene Gattung in der Abstammungslehre (Phylogenetik).
Dieser Regel folgen beide Ansätze. Allerdings fällt auf, dass nach der Analyse von Pyron & Burbrink (2009) auch das Konzept von Collins & Taggart (2008), welches auf der nicht so detaillierten und genauen Arbeit von Burbrink und Lawson (2007) basiert, inkonsistent wäre. Denn nach der neueren Analyse von Pyron & Burbrink (2009) hätten Collins & Taggart auch die Gattung Pituophis weiter aufspalten müssten, also analog zu der Verzweigung von Mintonius und Pantherophis.
Pyron & Burbrink (2009) verwenden hingegen die gemeinsamen Vorfahren der jeweils gesamten Gruppe (schwarze Punkte), wodurch sich lediglich zwei Gattungen, nämlich Pituophis und Pantherophis ergeben. Dieser Ansatz ist ebenso monophyletisch und daher ebenfalls richtig. In der Taxonomie gibt es zwei Arten von Wissenschaftlern. Die einen sind sogenannte "Splitter", die alles haarklein aufteilen (wie es Collins & Taggart 2008 getan haben) und die sogenannten "Lumper", die weniger fein, dafür aber praktikabel aufteilen, wie es Pyron & Burbrink (2009) getan haben. Es gibt kaum einen Grund Gattungen mit an sich wenigen Arten, die einen gemeinsamen Vorfahren haben (schwarze Punkte) noch weiter zu unterteilen, wie es Collins & Taggart (2008) getan haben. Daher hat sich der Ansatz von Pyron & Burbrink (2009) auch durchgesetzt. Ihre Einteilung wird auch von Crother und Co. (2011) akzeptiert und verwendet.
Demnach stellt sich die Gruppe der Kornnattern wie folgt dar:
- Pantherophis guttatus
- Pantherophis emoryi
- Pantherophis slowinskii
Die Erdnattern
Der ehemals aus sieben Unterarten bestehende Elaphe obsoleta-Komplex wurde 2000 von Burbrink, Lawson und Slowinski einer näheren genetischen Analyse unterzogen. Die beiden Unterarten E. o. deckerti und E. o williamsi wurden nicht von allen Autoren anerkannt, weswegen stellenweise nur von fünf Unterarten gesprochen wurde. Ihre Ergebnisse wiesen eindeutig darauf hin, dass sich die Unterarten in drei große Gruppen einordnen ließen. Die ganz im Osten beheimateten E. o. deckerti, E. o. rossaleni und E. o. quadrivittata, die zentral verbreiteten E. o. spiloides, E. o. williamsi und einige E. o. lindheimeri sowie die im Westen lebenden E. o. obsoleta und andere E. o. lindheimeri. Unabhängig davon bildeten die ebenfalls in die Untersuchung einbezogenen E. bairdi eine eigene Gruppe. Ebenfalls untersuchten sie die geographische Ausbreitung der Gruppen und stellten fest, dass diese aus drei verschiedenen Linien hervorgingen. Die westliche Linie stieß im Osten ihres Verbreitungsgebietes an den Mississippi der als geographische Barriere eine weitere Ausbreitung blockierte. Die zentrale Linie breitete sich östlich des Mississippi aus, ehe sie ihrerseits an ihrer östlichen Grenze auf das für sie unüberwindbare Appalachen Gebirge stieß. Südlich des Gebirges verhinderte der Appalachiola Fluss eine weitere Ausbreitung gen Osten. Die östliche Linie breitete sich dann zwischen dem Gebirge und der Ostküste Nordamerikas aus. Die Autoren merkten an, dass aufgrund dieser Ergebnisse der Status der Unterarten zweifelhaft sei und einer dringenden Überarbeitung bedürfe.
Im folgenden Jahr befasste Burbrink (2001) sich erneut mit dem obsoleta Komplex und zog aus diesen klaren Gruppierungen Konsequenzen. Er synonymisierte die Unterarten der jeweiligen Gruppe und erhob sie in den Status eigener Arten. Dazu musste er den ältesten verfügbaren Namen der jeweiligen Gruppe als Artnamen verwenden. Im Fall der östlichen Gruppe war dies E. alleghaniensis (Holbrook 1836), da Holbrook diesen in seinem Buch zwei Seiten vor quadrivittatus erwähnte und er somit als der älteste Name gilt. Die zentrale Gruppe wurde zu E. spiloides und die westliche Gruppe erhielt den Namen E. obsoleta. In den beiden letzten Gruppen fanden sich Exemplare von E. o. lindheimeri, was zu einer Aufteilung dieser Unterart führte. Das Verbreitungsgebiet von E. o. lindheimeri wird durch den Mississippi geteilt, so dass es keinen Genaustausch zwischen den westlich lebenden und den östlichen Tieren geben konnte. Auch die genetischen Untersuchungen bestätigten dies und so wurden die westlichen Exemplare E. obsoleta zugeordnet und die östlichen E. spiloides. Für die Terrarienhaltung bedeutet dies leider, dass alle derzeit erhältlichen lindheimeri (u.a. die beliebten Leuzisten) potenzielle Arthybriden sind solange sie keinen nachweislichen Stammbaum haben der den Fundort der Ausgangstiere und aller in der Linie beteiligten Tiere klärt.
Da Burbrink und Co. (2000) sowie Burbrink (2001) nur wenige Exemplare aus West Virginia berücksichtigt hatten, führte Mann (2007) eine umfassende morphologische Untersuchung von 114 Exemplaren unterschiedlicher Herkunft durch. Er wollte prüfen wie weit eine Zuordnung zu den von Burbrink (2001) eingeführten Arten möglich sei. Mann (2007) kam zu dem Ergebnis, dass sämtliche Erdnattern sehr variabel sind und eine rein morphologische Zuordnung nicht möglich sei. Daraufhin schlug er die Rückkehr zur alten Einteilung vor, da seiner Meinung nach diese Ergebnisse einen Artstatus nicht rechtfertigen würden. Seine Kritik hat ihre Berechtigung, da es nun in der Tat sehr schwierig ist Exemplare ohne Lokationsangabe rein auf Basis ihrer Morphologie sicher einer der drei Arten zuzuordnen. Interessanterweise zeigten Exemplare aus dem Verbreitungsgebiet von P. spiloides und P. alleghaniensis größere Ähnlichkeiten zueinander als zu P. bairdi und P. obsoletus, was sich auch in den genetischen Analysen von Burbrink und Co. (2000) ablesen lässt, da sich diese beiden Gruppen bereits seit längere Zeit durch den Mississippi getrennt voneinander entwickelt haben.
Die unterschiedlichen Ansichten der beiden Autoren (Burbrink und Mann) stammen von ihren unterschiedlichen Ansätzen. Burbrink verfolgt den sogenannten phylogenetischen Ansatz, welcher die Abstammungsgeschichte der einzelnen Arten anhand ihrer genetischen Unterschiede zu bestimmen versucht. Dabei erkennt er keine Unterarten an, sondern definiert alles, was sich deutlich genug voneinander unterscheidet, als eigenständige Art. Mann hingegen verwendet einen sogenannten morphometrischen Ansatz, welcher sich mit morphologischen Ähnlichkeiten und Unterschieden befasst. Populationen die nur schwach voneinander abweichen werden hier häufiger als Unterarten definiert. In der heutigen Wissenschaft werden genetische Analysen aber eher anerkannt als morpholgische Untersuchungen. Leider kann dies stellenweise dazu führen, dass manche Autoren Analysen durchführen ohne die Tiere je gesehen zu haben. Das Optimum läge in einer Mischung aus genetischen und morphologischen Untersuchungen, da rein genetische Analysen die ökologischen, morphologischen und biologischen Unterschiede eher selten berücksichtigen was oft zu Ergebnissen führt, die auf den ersten Blick unverständlich erscheinen.
Nach den Umgruppierungen durch Burbrink und Lawson (2007), Collins und Taggart (2008) sowie Pyron und Burbrink (2009) (siehe oben), stellt sich die Gruppe der Erdnattern wie folgt dar:
- Pantherophis bairdi
- Pantherophis alleghaniensis
- Pantherophis spiloides
- Pantherophis obsoletus
Die Fuchsnattern
Bei den Fuchsnattern gab es längere Zeit keine größeren Änderungen. Collins (1991) erhob die im Osten verbreitete Unterart Elaphe vulpina gloydi in den Artstatus und berief sich dabei auf die bereits von Conant (1940) angeführten geographischen und morphologischen Unterschiede zwischen den beiden Linien. Harding (1997) fügte ergänzend hinzu, dass beide Arten auch verschiedene ökologische Nischen in ihren jeweiligen Lebensräumen besetzen.
Allerdings wurde E. gloydi nicht von allen Autoren anerkannt. So taucht z.B. in Burbrinks Arbeiten stets E. vulpina auf, E. gloydi wird jedoch nicht erwähnt. Er begründet die Auslassung damit, dass Collins (1991) nur auf die von Conant (1940) ermittelten Daten zurückgegriffen hatte ohne diese um eigene und neue Daten zu erweitern. Seiner Ansicht nach entspricht dies nicht der üblichen Herangehensweise und wird demnach als nicht ausreichend belegt angesehen (Burbrink, pers. Mttlg.). Crother (2000) hingegen erwähnt E. gloydi in seiner Auflistung aller wissenschaftlichen und Trivialnamen als valide Art und weist zusätzlich auf eine unveröffentlichte Arbeit hin, welche den Artstatus von E. gloydi durch genetische Analysen bestätigt (Gardner, Crother und White).
Eine umfassende genetische Analyse von Crother und Co. (2011) führte zu dem Ergebnis, dass die ursprünglich von Conant (1940) angenommene Grenze zwischen den beiden Arten nicht die tatsächliche Verbreitungsgrenze darstellte. Es zeigten sich zwei deutliche Linien, die durch den Mississippi als natürliche Grenze voneinander getrennt wurden, was sich ebenso bereits bei den Kornnattern und Erdnattern gezeigt hatte. Von den 33 untersuchten Exemplaren wichen fünf von dieser deutlichen Zuordnung ab, woraus Crother und Co. (2011) folgerten, dass es einzelnen Exemplaren in jüngster Vergangenheit möglich war diese Barriere zu überwinden. Durch die Verschiebung der Verbreitungsgrenzen ergab sich aber nun die Notwendigkeit einer Umbenennung.
So wurde die Art P. gloydi aufgrund von Exemplaren beschrieben die sich im Verbreitungsgebiet weit östlich des Mississippi befanden. Die Art P. vulpinus wurde anhand von Exemplaren beschrieben die sich ebenfalls östlich des Mississippi befanden. Durch den Nachweis der genetischen Zusammengehörigkeit aller östlich lebenden Fuchsnattern hatte für diese Art der ältere Name Vorrang: In diesem Fall P. vulpinus (Baird und Girard, 1853). Damit blieben aber alle westlich des Mississippi lebenden Fuchsnattern unbenannt, da hier kein Referenzexemplar (Holotypus) bekannt war. Es gab noch einen weiteren Namen für die Fuchsnattern, E. rubriceps. Dieser wurde von Duméril und Bibron (1854) eingeführt. Da der von ihnen beschriebene Holotypus aber leider verloren gegangen war konnte man ihn nicht mehr in die genetische Analyse mit einbeziehen um ihn einer der beiden Linien zuzuordnen. Conant (1940) synonymisierte E. rubriceps mit E. vulpina, da die Beschreibung des Holotypus mehr auf diese Art zutraf. Mangels Nachvollziehbarkeit der genauen Herkunft des von Duméril und Bibron (1854) beschriebenen Exemplars sehen Crother und Co. (2011) den Namen E. rubriceps als nicht verfügbar an. Demnach gab es keinen gültigen Namen für die westliche Linie, so dass Crother und Co. (2011) Pantherophis ramspotti als Namen für diese Linie einführten.
Anzumerken ist hierbei eine wesentliche Änderung bei den Trivialnamen. Während man bislang P. vulpinus als westliche Fuchsnatter und P. gloydi als östliche Fuchsnatter bezeichnete, ist nach der Änderung von Crother und Co. (2011) P. vulpinus die östliche Fuchsnatter während P. ramspotti die westliche Fuchsnatter ist.
Nach den Umgruppierungen durch Burbrink und Lawson (2007), Collins und Taggart (2008) und Pyron und Burbrink (2009) (siehe oben) sowie nach der Neubeschreibung durch Crother und Co. (2011) stellt sich die Gruppe der Fuchsnattern wie folgt dar:
- Pantherophis vulpinus
- Pantherophis ramspotti
Quellen
BURBRINK, FRANK T. (2001) Systematics of the North American Rat Snake Complex (Elaphe obsoleta). Herpetological Monographs 15:1–53.
BURBRINK, FRANK T (2002) Phylogeographic analysis of the cornsnake (Elaphe guttata) complex as inferred from maximum likelihood and Bayesian analyses. Molecular Phylogenetics and Evolution 25:465–476.
BURBRINK, FRANK T., AND ROBIN LAWSON (2007) How and when did Old World rat snakes disperse into the New World? Molecular Phylogenetics and Evolution 43:173–189.
BURBRINK, FRANK T., LAWSON, ROBIN, AND JOSEPH. B. SLOWINSKI (2000) Molecular phylogeography of the North American rat snake (Elaphe obsoleta): A critique of the subspecies concept. Evolution 54(6):2107–2114.
COLLINS, JOSEPH T. (1991) Viewpoint: A new taxonomic arrangement for some North American amphibians and reptiles. Herpetological Review 22(2): 42–43.
COLLINS, JOSEPH T. (2010) An Accurate Accounting of an Inaccurate Snake Taxonomy. Journal of Kansas Herpetology 36: 14–15.
COLLINS, JOSEPH T. UND TRAVIS W. TAGGART (2008) An Alternative Classification of the New World Rat Snakes (Genus Pantherophis [Reptilia: Squamata: Colubridae]). Journal of Kansas Herpetology 26:16–18.
CONANT, ROGER (1940) A New Subspecies of the Fox Snake, Elaphe vulpine Baird and Girard. Herpetologica 2(1): 1–15.
CROTHER, BRIAN I. (2000) Scientific and Standard English Names of Amphibians and Reptiles of North America North of Mexico, With Comments Regarding Confidence in Our Understanding, 6. Auflage 2000, SSAR.
CROTHER, BRIAN I., WHITE, MARY E., SAVAGE, JAY. M, ECKSTUT, MALLORY E., GRAHAM, MATTHEW R., UND DAVID W. GARDNER (2011) A Reevaluation of the Status of the Fox snakes Pantherophis gloydi Conant and P. vulpinus Baird and Girard (Lepidosauria). International Scholarly Research Network (ISRN) Zoology, 1–15.
HARDING, J. H. (1997) Amphibians and Reptiles of the Great Lakes Region. University of Michigan Press, 378 Seiten.
MANN, ADAM M. (2007) A Taxonomic Investigation of the Black Ratsnake, Elaphe o. obsoleta (Say) [Reptilia, Squamata, Colubridae], in West Virginia using Morphometric Analyses. Master-Arbeit, Marshall Universität, 2007, 101 Seiten.
PYRON, R. ALEXANDER UND FRANK T. BURBRINK (2009) Neogene diversification and taxonomic stability in the snake tribe Lampropeltini (Serpentes: Colubridae). Molecular Phylogenetics and Evolution 52:524–529.
UTIGER, URS, HELFENBERGER, NOTKER, SCHÄTTI, BEAT, SCHMIDT, CATHERINE, RUF, MARKUS UND VINCENT ZISWILER (2002) Molecular Systematics and Phylogeny of old and new world rat snakes, Elaphe AUCT., and related genera (Reptilia, Squamata, Colurbidae). Russian Journal of Herpetology 9(2):105–124.
Hinweis
Leider muss ich an dieser Stelle auch darauf hinweisen, dass jegliche Weiterverbreitung (auch auszugsweise) ohne mein schriftliches Einverständnis zu unterlassen ist. Wer diesen Artikel auf seiner Homepage oder anderswo verbreiten möchte, nutzt bitte den Link zu dieser Seite! Danke.
Artikel erstellt von Sabrina Schleip, 2012
Original erschienen ist mein Artikel 2012 im alten Farbvarianten.de-Forum.